Preliminary report on taxonomical position of Vavilovia formosa (Stev.) Fed. evidenced from morphological and molecular data
Sinjushin, A.A.,
Demidenko, N.V. and M.V. Lomonosov Moscow State University, Moscow, Russia
Gostimskii, S.A.
The Fabeae Rchb. (syn. Vicieae Adans.) tribe includes numerous representatives of special agricultural value and has; therefore, received considerable attention. Despite (or because of) this, there are certain problems concerning its systematics and phylogeny. One of the most problematic groups within the tribe is the monotypic genus Vavilovia Fed. It includes very small perennial plants inhabiting the highlands of the Caucasus, Turkey, Lebanon and Iran. Although almost two centuries have passed since the first description of this plant was reported by C. Steven (more detailed information on the history of its taxonomical status is presented in (1)), its status remains unresolved. Previously it was included in the genera Orobus, Lathyrus, Pisum, or treated as a separate genus, Alophotropis Grossh. or Vavilovia Fed. The most recent monographs on the family tend to describe its status as a monotypic genus including a single species, Vavilovia formosa (Stev.) Fed. (2).
Vavilovia inhabits very specific areas and all efforts to maintain the plant beyond its natural habitat failed or presented significant difficulties (3, 4). Moreover, it is an endangered species with disrupted distribution areas, some of which are in danger of being overgrazed (5). Probably due to paucity of material, the genus remains poorly studied and until now few efforts have been made to use molecular data to resolve its taxonomical status. The only available work in which some features of the chloroplast genome were studied was by Jansen et al. (6), but this sheds no light on the position of V. formosa in the tribe. Strong relationship between Vavilovia and Pisum was demonstrated basing on sequences of both cpDNA and nrDNA regions and discussed in (7) and (8). Some other similarities such as susceptibility to pea-specialized fungal infections, chromosome number and successful crosses between Pisum and Vavilovia, etc., also indicated a strong relationship. It should be noted; however, that the possibility of mutual hybridization between listed genera (4) can hardly be interpreted as a feature of relationship. For example, some reports on crosses between P. sativum and Vicia faba exist (9, see also 10), while no success was reached in crosses between Pisum and Lathyrus. The same can be concluded based on chromosome number, where 2n=14 remains constant for all genera within Fabeae (with rare exceptions) together with numerous related legumes (such as tribe Loteae DC.). Some differences in chromosome morphology between these genera were revealed in (11).
This work was aimed at clarification of the status of Vavilovia within the tribe Fabeae with special reference to its relation with Pisum. This paper represents the preliminary results of the first stages of our investigation.
Materials and Methods
Plant material
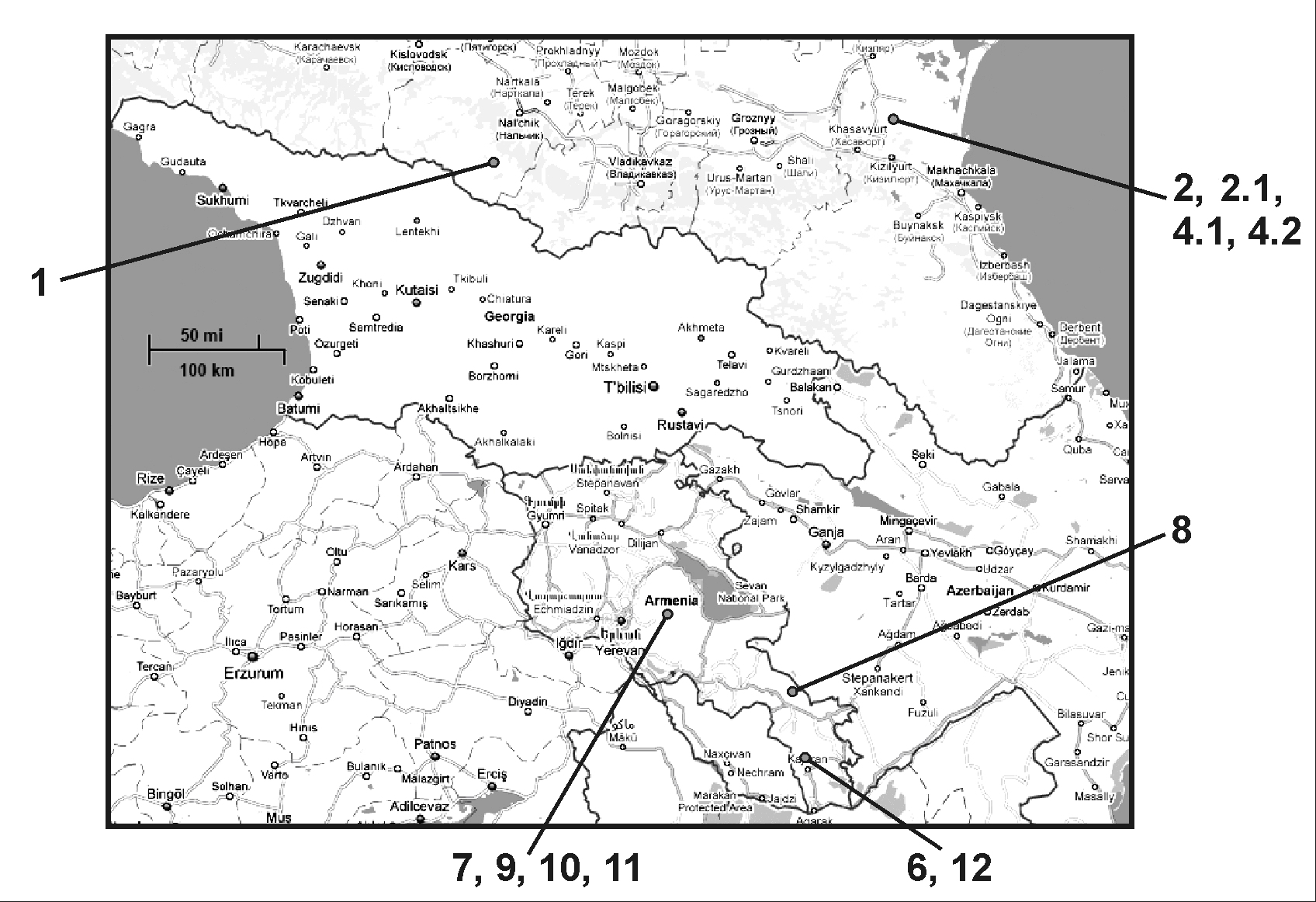
Herbarium specimens of Vavilovia formosa from Moscow State University Herbarium (MW) and the Institute of Botany of the National Academy of Sciences (Republic of Armenia) were used as the material for this study. These samples represent material from different parts of the area of habitat of studied specimens in the Caucasus (Fig. 1); a total of 14 accessions were studied. Six Pisum sativum L. ssp. sativum cultivars from different world regions, four wild P. sativum subspecies and one P. fulvum Sibth. & Smith accession from the Genetics Department of Moscow State University and John Innes Center (Norwich, United Kingdom) Pisum germplasm collections were used for comparison. Eleven morphologically contrasting Lathyrus species were analyzed; L. sativus L., L. tingitanus L., L. odoratus L., L. belinensis N. Maxted & Goyder, L. chloranthus Boiss. & Balansa, L. ochrus DC., L. aleuticus (Greene) Pobed., L. nissolia L., L. vernus Bernh., L. venetus Rouy, and L. aureus (Steven) Bornm. (the three latter species were formerly placed in genus Orobus but are now treated as part of Lathyrus). Seeds of the first six species were kindly provided by Dr. G.D. Levko (All-Russian Research Institute of Breeding and Seed Production of Vegetables, Lesnoi Gorodok, Russian Federation). All listed material was used for both morphological and molecular analysis.
Figure 1. Map depicting locations of sample collection. The site of collection of accession number 5 was unclear.
DNA extraction, PCR conditions and sequencing
DNA was extracted from freshly collected leaflets and herbarium specimens using a modified CTAB procedure (12). For homogenization of herbarium material, glass powder was added to the plant sample. PCR was carried out in a MC2+ Thermal Cycler (DNA Technology, Russia) according to a protocol described in earlier work (13). At this stage, only RAPD (Randomly Amplified Polymorphic DNA) markers were used to characterize polymorphism while usage of other marker types is in progress. The following primers were used: V (5Æ-gacagtagca-3Æ), VI (5Æ-cttggatgga-3Æ), V03 (5Æ-ctccctgcaa-3Æ), B474 (5Æ-aggcgggaac-3Æ), D6 (5Æ-acctgaacgg-3Æ), F12 (5Æ-acgctaccag-3Æ), R11 (5Æ-gtagccgtct-3Æ), Q06 (5Æ-gagcgccttc-3Æ), QR2 (5Æ-cggccactgt-3Æ), AE07 (5Æ-gtgtcagtgg-3Æ), AE13 (5Æ-tgtggactgg-3Æ), Leb10 (5Æ-agccgcagct-3Æ) (Syntol, Russian Federation). A total of 194 polymorphic bands were obtained.
The internal transcribed spacer region (ITS1-5.8 rRNA-ITS2) was amplified using primers flanking the ITS1-5.8 rRNA-ITS2 region of the nuclear genome. Primer sequences are available from (14) and (15) for forward and reverse primers, respectively. The fragment obtained from the sample designated Vf2.1 (Fig. 1) was sequenced using an automatic sequencer (ABI Prism 3100-Avant Genetic Analyzer) in the ōGenomeö Center (Institute of Molecular Biology of Russian Academy of Sciences) and used for further study.
Tree construction
Taxonomical analysis was performed by comparing obtained sequences with the corresponding region of all genera of the Fabeae tribe (Pisum, Lathyrus, Vicia, Lens) arbitrarily chosen from GenBank (http://www.ncbi.nlm.nih.gov/Genbank/index.html) database. Two Cicer (Cicereae) sequences were chosen as an outgroup. Phylogenetic analyses were conducted using the MEGA4 software (16). Data obtained from RAPD analysis was processed with usage of SplitsTree 4.10 software. The distances were calculated according to formula of L. Dice (17).
Figure 2. Morphology of Vavilovia formosa. Frame indicates the leaf of Armenian samples.
Results and Discussion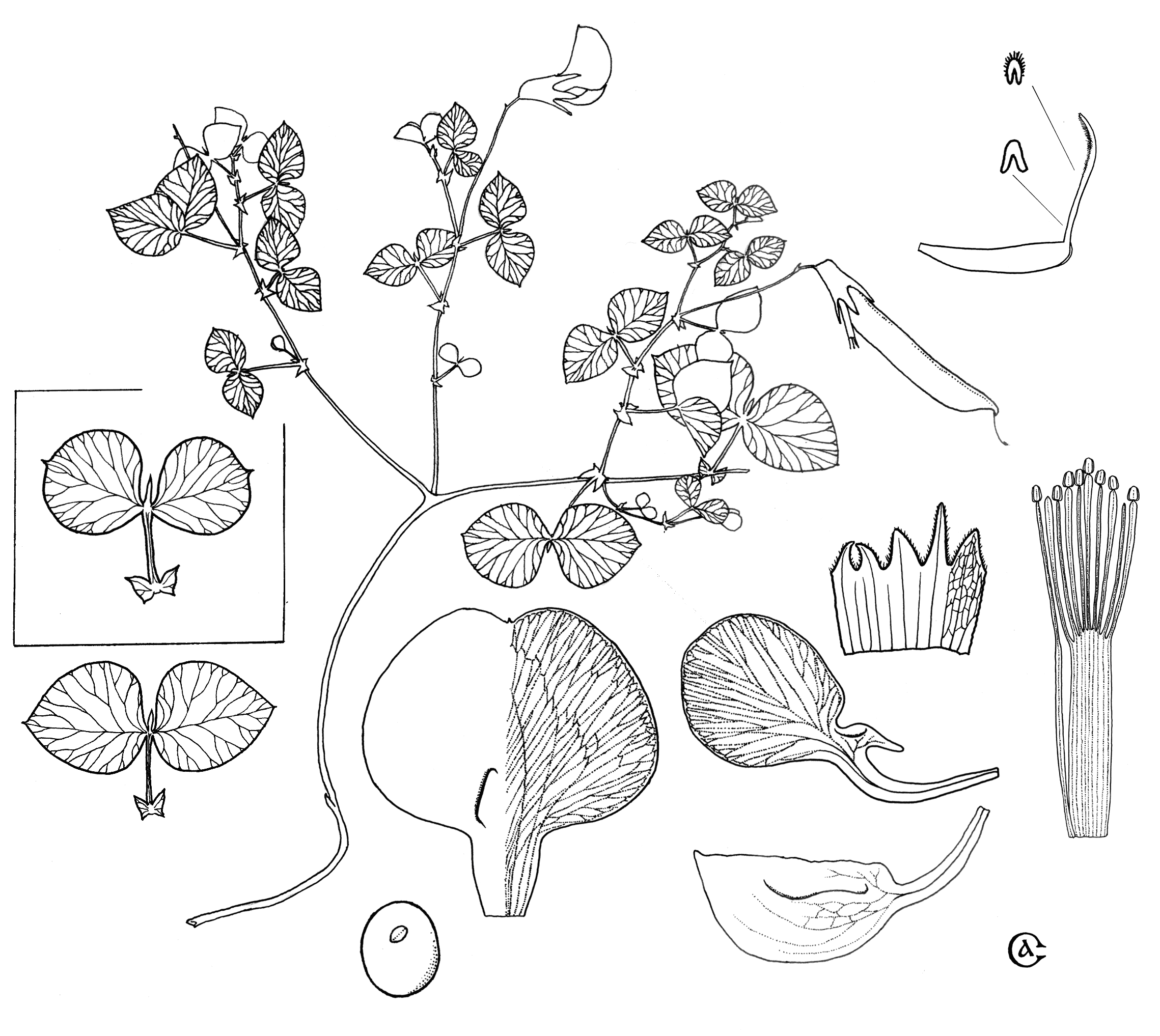
Morphological features of Vavilovia
The Vavilovia plants are characterized with very specific morphology (Fig. 2). Some features were found which distinguish them from all studied Pisum accessions. These features include: perennial life form (Pisum annual) with long creeping rhizomes enabling vegetative propagation; narrow sepals covered with simple trichomes (glabrous and broad with overlapping margins in Pisum); leaf rachis ending with cusp (always tendril in Pisum); small stipules (of the same size as leaflets or larger in Pisum); fluted leaf rachis cross-section (rounded in Pisum); supervolute ptyxis (plicate in Pisum, this trait is reported as one of high significance by F. Kupicha (18)). All the listed characters can be found in Lathyrus species, but their interpretation is ambiguous. The life form (annual or perennial) is not used even to separate sections in Lathyrus. Leaf features are known to be very variable in ontogeny of legumes, and the first pea leaves are always characterized with inversely-ovate leaflets, fluted rachises ending with short process or even lacking any specialized terminus, and even small stipules (P. fulvum). Actually, Vavilovia leaves combine some features which can be found in juvenile forms of Pisum species. The calyx of the latter also possess ciliate margins before anthesis. The only feature of special significance is ptyxis which was reported to be altered in mutational variation of Vicia faba (18).
The ovary of the Pisum flower is glabrous (C. Gunn and J. Kluve report glandular surface for some accessions (10)), while Lathyrus species exhibit both glabrous and pubescent (simple trichomes, nectariferous glands or combination of two types (10, original observations)) ovaries. Vavilovia samples we studied possessed a combination of simple trichomes and few-celled glands on the surface of the ovaries.
Some characters which distinguish Vavilovia from all studied Lathyrus species (and hence drawing with peas together) include; elliptic seed hilum (linear or rarely elliptic in Lathyrus); absence of pubescence on all organs except the calyx; and dilated filaments of outer whorl stamens (as they are in Pisum; somewhat dilated filaments are also found in L. maritimus).
It should be noted that these characters had never been found altered even in mutational variation of Pisum and hence can be referred to as strongly constant for genus identity. Standard (vexillum) petal shape of Vavilovia also resembles that of Pisum rather than Lathyrus.
Two traits were cited to separate Vavilovia from Pisum, viz. pistil groove gaping only at the base (at base and apex in Pisum) and non-cristate keel (cristate in pea) (see (10); these authors reject these differences). Pistils appear to be grooved in the same way in both genera, as do other pistil features such as hair pattern (adaxial). As for keel crest, this feature is difficult to analyze on herbarium material and needs more detailed investigations.
At least two features distinguish Vavilovia from all other studied species. These are leathery leaflets and specific leaflet shape. In all studied plants, proximal leaflet half (i.e. one directed to leaf basement) is wider than the distal (directed to leaf rachis tip) or has almost the same width. In all Vavilovia plants distal half is notably wider than proximal.
The Vavilovia specimens are not completely uniform in the sample studied. All Armenian plants differed from the rest in having leaflets with keel-like bases (rounded base in other specimens, Fig. 2). This feature was used by A.A. Grossheim (19) to separate two different species within Alophotropis (=Vavilovia), namely A. formosa and A. aucheri, but in our opinion the whole genus should be treated as monotypic (at least basing on available material), probably with the contrast in forms interpreted as subspecies. The morphologically distinct forms of Pisum formosum (= V. formosa) were treated as separate varieties by L. Govorov (20), viz. var. typicum Gov. and var. microphyllum Ser.
DNA analysis
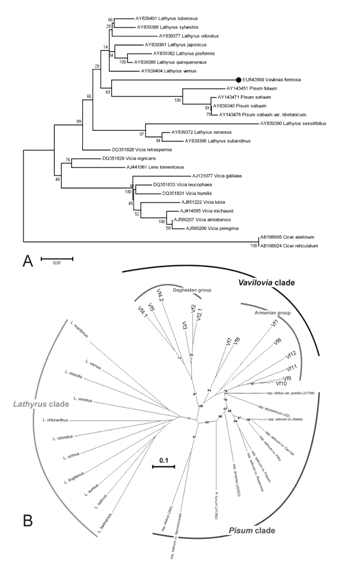
Analysis of the RAPD data revealed three clades (Fig. 3B). The Lathyrus accessions were strongly different from other accessions and formed a single clade, but differences between them provide no opportunity to uncover any interrelations within the genus. Even ōoroboidö species did not form a single group thus providing evidence that Lathyrus sections cannot be keyed out with this method (or at least with such resolution). Notably, variation between representatives of single genus Lathyrus are much more expressed than between forms interpreted as separate genera, Pisum and Vavilovia. Vavilovia accessions were distinct from other samples and were subdivided into two groups according to geographical position of sites of collection, Daghestan and Armenian groups.
The comparison of nucleotide sequences of ITS region of representatives of all genera comprising Fabeae tribe exhibited some controversy to morphological features and revealed that Vavilovia demonstrates relation to Pisum rather than to other genera (Fig. 3A). However, it can be concluded that V. formosa is more distant from type pea subspecies (P. sativum ssp. sativum) than even P. fulvum. The differences between Armenian and Daghestan specimens can be interpreted as corresponding to subspecies rather to distinct species or varieties.
Obviously, more detailed investigations on this point are needed but preliminary data presented here supports the similarity of Pisum and Vavilovia. In our opinion, the latter genus can be even treated as part of Pisum sativum L., i.e. as Pisum formosum (Stev.) Alef. The individuality of discussed species can probably serve as a basis for separation of P. formosum into a monotypic section. This idea was earlier proposed by some investigators (see (2)); for example, Pisum sativum L. was treated as subdivided into sections Alophotropis Jaub. et Spach (including Vavilovia) and Lophotropis Jaub. et Spach (including Pisum sativum Str.).
Figure 3. Dendrograms of Vavilovia samples and related species based on sequences of ITS1-5.8 rRNA-ITS2 region of the nuclear genome (A) and RAPD analysis (B). The percentage of replicate trees in which the associated taxa clustered together in the bootstrap test (100 replicates for A, 1000 for B) is shown next to the branches. Numbers of V. formosa specimens (ōVfö series) correspond to those from Figure 1. Vavilovia accessions are denoted with a deimal point in A.
Acknowledgements: Authors cordially express their gratitude to Dr. J.A. Akopian (Institute of Botany of the National Academy of Sciences, Republic of Armenia) for providing herbarium specimens from the territory of Armenia; Dr. M. Ambrose (John Innes Centre, Great Britain) and Dr. G.D. Levko (All-Russian Research Institute of Breeding and Seed Production of Vegetables, Russian Federation) for kindly providing seeds of Pisum and Lathyrus species. Authors are also indebted to Dr. F.A. Konovalov (N.I. Vavilov Institute of General Genetics) and N.A. Poyarkov (Biological Faculty of Moscow State University) for help in computer analysis. The work was partially supported by the Russian Foundation for Basic Research (grant no. 07-04-00652) and the ōLeading Scientific Institutionsö program.
1. Makasheva, R.K., Drozd, A.M., Adamova, O.P. and A.A Golubev. 1973. Bull. Appl. Bot., Genet. Pl. Breed. 51:44-56 (In Russian).
2. Lock, J.M. and Maxted, N. 2005. In: G. Lewis, B. Schrire, B. Mackinder and M. Lock (eds.). Legumes of the world. Kew. p. 504-509.
3. Cooper, S.R. and Cadger, C.A. 1990. Pisum Newslett. 22:5.
4. Golubev, A.A. 1990. Bull. Appl. Bot., Genet. Pl. Breed. 135:67-75 (In Russian).
5. Akopian, J.A. and Gabrielyan, I.G. 2008. Crop Wild Relative 6:26-27.
6. Jansen, R.K., Wojciechowski, M.F., Sanniyasi, E., Lee, S.-B. and Daniell, H. 2008. Mol. Phylogenet. Evol. 48:1204-1217.
7. Kenicer, G., Sm²kal, P., Vishnyakova, M. and Mikić, A. 2008. Grain Legumes 51:8.
8. Sm²kal, P., Kenicer, G.and Mikić, A. 2009. Book of Abstracts of IV Congress of the Serbian Genet. Soc. p. 166.
9. Novikov, V.V. 1940. Yarovizatsiya 3:156-167 (In Russian).
10. Gunn, C.R. and Kluve, J. 1976. Taxon 25:563-575.
11. Abramova, L.I. 1971. Bull. Appl. Bot., Genet. Pl. Breed. 45:240-243 (In Russian).
12. Torres, A.M., Weeden, N.F. and Martin, A. 1993. Theor. Appl. Genet. 85:937-945.
13. Konovalov, F., Toshchakova, E. and Gostimsky, S. 2005. Cell. Mol. Biol. Lett. 10:163-171.
14. Hsiao, C., Chatterton, N.J., Asay, K.H. and Jensen, K.B.. 1994. Genome 37:112¢120.
15. White, T.J., Bruns, T., Lee, S. and Taylor, J.W. 1990. In: M.A. Innis, D.H. Gelfand, J.J. Sninsky and T.J. White, (eds.). PCR Protocols: A Guide to Methods and Applications. New York. p. 315¢322.
16. Tamura, K., Dudley J., Nei, M. and Kumar, S. 2007. Mol. Biol. Evol. 24:1596-1599.
17. Dice, L.R. 1945. Ecology 26:297-302.
18. Kupicha, F. 1981. Vicieae. In: R.M. Polhill and P.H. Raven (eds.). Advances in legume systematics. 1:377-381.
19. Grossheim, A.A. 1949. Identification of plants of the Caucasus. Moscow, p. 162.
20. Govorov, L.I. 1937. In: N.I. Vavilov and E.V. Wulff (eds.). Flora of cultivated plants. IV. Grain Leguminosae. Moscow, Leningrad. p. 248-249.