|
PNL |
Volume 22 |
1990 |
RESEARCH REPORTS |
pages 48-49 |
INTERNODE LENGTH IN PISUM. THE GENOTYPE OF GOTTSCHALK'S MUTANT 84B
|
Reid, J.B.
|
Department of Plant Science, University of Tasmania Hobart, Tasmania 7001, Australia |
Internode length mutants in peas have proved extremely valuable as research tools in plant physiology (4). For example, Gottschalk's dwarf mutants M26 and M66 have provided the recessive mutations ls and led, which, when homozygous, produce plants that are deficient in GA1 (5,7). These mutants have allowed it to be shown that a clear quantitative relationship occurs between the levels of GA1 and elongation (7). M84B is a further dwarf mutant produced by Professor Gottschalk (Institut f�r Genetik, Bonn) from cv Dippes Gelbe Viktoria (DGV) by X-irradiation (Gottschalk, pers. comm.) and it may also be of use in physiological research. The parental cv DGV possesses the internode length genotype le La cryc Lm Na Lh Ls Lk Lka Lkb Lw Lv (2) and it has the longest internodes of any dwarf variety in our collection at Hobart.
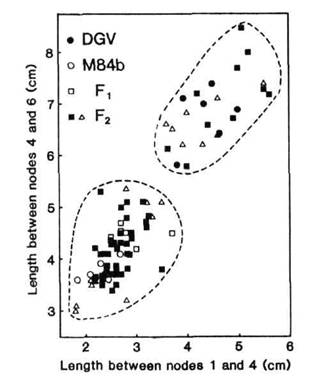
The nature of the mutation in M84B was first examined by crossing M84B to cv DGV. The F1 possessed marginally longer internodes than the M84B parent and was substantially shorter than the parental cv DGV (Fig. 1). In the F2 a relatively clear segregation into 50 short and 18 long plants was obtained (Fig. 1). This is in good agreement with a 3:1 segregation (c21 = 0.08) suggesting that M84B differs from DGV by possessing a dominant mutation. Although a clear separation across families did not occur in F3, analysis of F3 data within families supports this conclusion since progeny from 10 short parents segregated to give 52 short and 16 long plants (c21 testing 5:1 = 2.9) while progeny from 8 long F2 parents bred true (32 plants). If it is assumed that the genetic background is the same in DGV and M84B the degree of dominance can be obtained directly from the F1 data and, based on the length between nodes 1 and 6, is 0.4. F3 data support this lack of complete dominance.
Crosses of both DGV and N84B were made to the Hobart dwarf line 61a (le la Cry Lm Na Ls Lh Lw Lk Lka Lkb Lv; 6). The F2 of cross 61a x M84B yielded no clear segregation for internode length (Fig. 2a) while the F2 of cross DGV x 61a segregated to produce 88 dwarf (La and/or Cry) and 8 cryptodwarf (la cryc) progeny (Fig. 2b). This latter segregation is in agreement with the expected 15:1 ratio (c21 = 0.4). The cryptodwarf plants were separated from the dwarf segregates by the stem length between nodes 1 and 4. The dominant mutation in M84B therefore may be at the previously established locus, Cry (1,3,6). The allele cryc in DGV has probably been replaced in M84B by an allele comparable in effect to Cry. However, whether this is a back-mutation to the wild-type allele at the DNA level remains unknown.
de Haan, H. 1927. Genetica 9:481-497.
Murfet, I.C. 1988. Ann. Bot. 61:331-345.
Rasmusson, J. 1927. Hereditas 10:1-150.
Reid, J.B. 1990. In Plant Growth Substances 1988, Eds R.P. Pharis and S.B. Rood, Springer-Verlag, Heidelberg, pp 74-83.
Reid, J.B. and W.C. Potts. 1986. Physiol. Plant. 66:417-426.
Reid, J.B., I.C. Murfet and W.C. Potts. 1983. J. Exp. Bot. 34:349-364.
Ross, J.J., J.B. Reid, P. Gaskin and J. MacMillan. 1989. Physiol. Plant. 76:173-176.
Fig. 1. Stem length between nodes 1 and 4 versus stem length between nodes 4 and 6 for the F1 (□), F2 (■, D) and parents of cross DGV (●) x N84B (○). F3 progenies were grown from the F2 plants indicated (D). The photoperiod was 18 h.
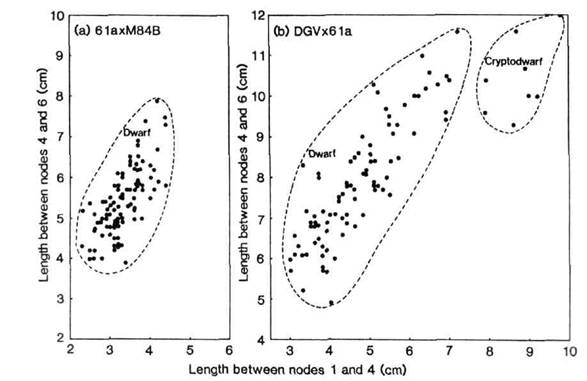
Fig. 2. Stem length between nodes 1 and 4 versus stem length between nodes 4 and 6 for the F2 of crosses 61a x M84B (Fig. 2a) and DGV x 61a (Fig. 2b).