Dominant alleles of the locus Tl
may differ in their strength
Bogdanova,
V.S., Kosterin O.E. and
Institute of
Cytology and Genetics of Russian Academy of Sciences
Berdnikov,
V.A.
Acad. Lavrentiev ave. 10, Novosibirsk 630090, Russia
The Tendril-less (Tl)
gene of Pisum sativum participates in controlling development of the
compound leaf and is located in the linkage group V. A number of recessive
alleles have been described. Homozygotes tlw,
tlpet condition formation
of leaflets in place of tendrils (2). Heterozygotes
exhibit an intermediate structure that can be called 'flat tendril', that is
tendril with a very narrow leaf blade. Another recessive allele tlx
is supposed to be a small deletion (3), and only was observed in hemizygote
which appears quite similar to the heterozygote Tl/tlw.
However, little is known of variants of dominant alleles. Here we show that
probably there exist at least two Tl
alleles that differ in their strength, that is, visible expression in
heterozygote against tlw.
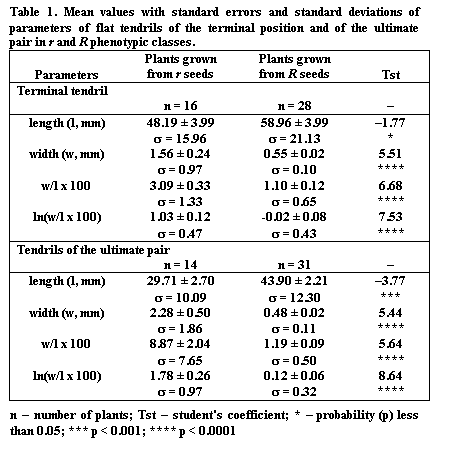 In 1997 we performed a cross of the lines Delta and Sprint-Vologda as a
part of investigation of the line Delta. The lines are described in (1), an
important point for the present study is that Delta is heterozygous (r Tl/R tlx) and Sprint-Vologda is homozygous (R
Tl/R Tl), the genes R and Tl being closely linked (4). For convenience, we will designate the Tl
allele originating from the line Delta as TlD
and that from the Sprint-Vologda line as TlSV.
Thus, in the F1 from the cross Delta x Sprint-Vologda there appear
plants with flat tendrils heterozygous for Tl
(R tlx/R TlSV)
and plants with round tendrils homozygous for Tl (r TlD /R TlSV).
Plants of the latter class were test-crossed as a pollen parent with the line
WL1018 (r tlw). We obtained
16 wrinkled (r tlw/r
TlD) and 49 round (r tlw/R
TlSV) seeds. The ratio of r
to R seeds differed significantly from
the expected 1:1, perhaps due to peculiarities of the line Delta (1). All plants
resulting from this test-cross expressed flat tendrils. Visual comparison of
flat tendril width in these two classes suggested a significant difference. We
therefore measured length and width of flat tendrils in the leaves at node 8 in
individual plants. The results are presented in Table 1.
In 1997 we performed a cross of the lines Delta and Sprint-Vologda as a
part of investigation of the line Delta. The lines are described in (1), an
important point for the present study is that Delta is heterozygous (r Tl/R tlx) and Sprint-Vologda is homozygous (R
Tl/R Tl), the genes R and Tl being closely linked (4). For convenience, we will designate the Tl
allele originating from the line Delta as TlD
and that from the Sprint-Vologda line as TlSV.
Thus, in the F1 from the cross Delta x Sprint-Vologda there appear
plants with flat tendrils heterozygous for Tl
(R tlx/R TlSV)
and plants with round tendrils homozygous for Tl (r TlD /R TlSV).
Plants of the latter class were test-crossed as a pollen parent with the line
WL1018 (r tlw). We obtained
16 wrinkled (r tlw/r
TlD) and 49 round (r tlw/R
TlSV) seeds. The ratio of r
to R seeds differed significantly from
the expected 1:1, perhaps due to peculiarities of the line Delta (1). All plants
resulting from this test-cross expressed flat tendrils. Visual comparison of
flat tendril width in these two classes suggested a significant difference. We
therefore measured length and width of flat tendrils in the leaves at node 8 in
individual plants. The results are presented in Table 1.
The dimensions of the flat tendrils, particularly their width:length
ratio, in two phenotypic classes, differ signi-ficantly.
As one of the pheno-typic classes has a much higher variance for this
character, it is reasonable to perform logarith-mic transformation.
After this procedure, the difference between plants carrying TlD
and TlSV alleles (more precisely, grown from r
and R seeds) becomes more evident.
Distri-butions of the logarithm of the
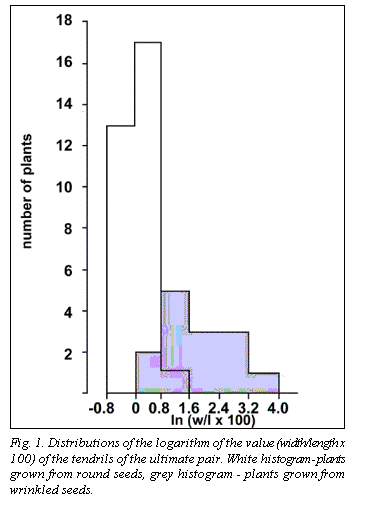 value (width/length
x 100) for the tendrils of the ultimate pair are shown in Fig.1. The difference
observed indicates that a factor controlling the width of flat tendrils
co-segregates with the alleles of R
locus. It could be some modifier closely linked to R and, therefore, to Tl.
However, we believe the effect is more likely due to the Tl gene itself. The distributions overlap to some extent, probably
due to high values of the variance or, possibly, misclassification of Tl-classes
due to rare crossing-over between r
and Tl (or between r and the
hypothetic modifer). These results allow us to propose that the two wild-type Tl
alleles studied in this work differ in their strength manifested in the width of flat tendrils of the
heterozygote Tl/tlw. The TlD
allele originating from the line Delta may be termed ōweakö, and the TlSV allele that comes from the line Sprint-Vologda may
be regarded as ōstrongö.
value (width/length
x 100) for the tendrils of the ultimate pair are shown in Fig.1. The difference
observed indicates that a factor controlling the width of flat tendrils
co-segregates with the alleles of R
locus. It could be some modifier closely linked to R and, therefore, to Tl.
However, we believe the effect is more likely due to the Tl gene itself. The distributions overlap to some extent, probably
due to high values of the variance or, possibly, misclassification of Tl-classes
due to rare crossing-over between r
and Tl (or between r and the
hypothetic modifer). These results allow us to propose that the two wild-type Tl
alleles studied in this work differ in their strength manifested in the width of flat tendrils of the
heterozygote Tl/tlw. The TlD
allele originating from the line Delta may be termed ōweakö, and the TlSV allele that comes from the line Sprint-Vologda may
be regarded as ōstrongö.
Acknowledgement:
The work was partly supported by the Russian State Program æRussian Fund for
Fundamental ResearchÆ, Grant No 99-04-49-970.
1. Berdnikov,
V.A., GorelÆ, F.L., Bogdanova, V.S., Kosterin, O.E., Trusov, Y.A., Rozov, S.M.
1999. Genet. Res. 73: 93-109.
2. Blixt,
S. 1972. Agri Hortique Genet. 30: 1-293.
3. GorelÆ,
F.L., Berdnikov, V.A., Temnykh, S.V. 1994.
Pisum Genet.. 26: 16-17.
4. Weeden,
N.F., Ellis, T.H.N., Timmerman-Vaughan, G.M., Swiecicki, W.K., Rozov, S.M.,
Berdnikov, V.A. 1998.
Pisum Genet. 30: 1-4.