|
PNL |
Volume 22 |
1990 |
RESEARCH REPORTS |
pages 6-11 |
INTERNODE LENGTH IN PISUM. GENOTYPES la cryS lka AND la cryS lkb AND THE PHENOTYPE SLENDER SEMI-ERECTOIDES
|
Cramp, R.E. and Reid, J.B.* |
Department of Plant Science, University of Tasmania Hobart, Tasmania 7001, Australia |
*Corresponding author. Study supported in part by an ARC grant.
The mutations lka (NGB5865) and lkb (NGB5862) both result in plants with a short, semi-erectoides phenotype (3). The response of both mutants to applied gibberellin A1 (GA1) is reduced in comparison with that of the tall, wild-type initial line (cv Torsdag) and thus they were classified as GA-insensitive mutants (3). The slender gene combination, la cryS, has also been shown to influence aspects of GA sensitivity even though the plants are elongated compared with wild type plants (2). It is therefore of interest to examine the interaction of the gene combination la cryS with both lka and lkb.
Mutations lka and lkb were induced in cv Torsdag by Dr K.K. Sidorova at Novosibirsk, USSR. The slender line used for the crosses, L197 (Le la cry5 Na Lh Lk Lka Lkb Lm Ls Lw Lv), was selected from a cross between NGB1766 (nana, Le La Cry na Lh Lk Lka Lkb Lm Ls Lw Lv) and L133 (slender, le la cryS Na Lh Lk Lka Lkb Lm Ls Lw Lv). Growing conditions were the same as those detailed by Reid and Ross (3). Counting of nodes began from the first scale leaf as node 1.
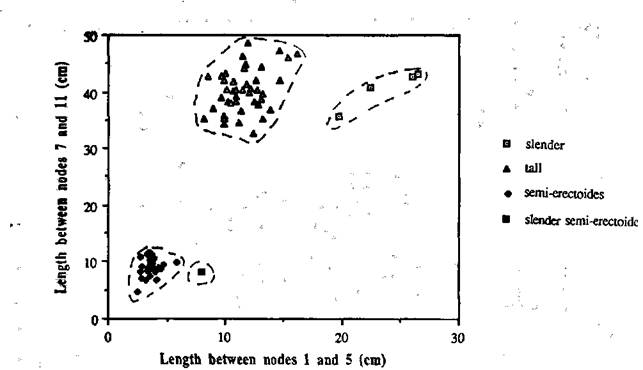
The F1 plants of crosses L197 x NGB5862 and L197 x NGB5865 possessed the wild type tall phenotype. The F2 of cross L197 x NGB5862 gave a clear segregation into 43 tall, 34 semi-erectoides, and 4 slender plants when the length between nodes 1 and 5 was compared with that between nodes 7 and 11 (Fig. 1). This segregation is not in accord with the expected ratio of 45 tall (La and/or Cry, Lkb) : 16 semi-erectoides (La and/or Cry, lkb and la cryS lkb) : 3 slender (la cryS Lkb) (c22 for 45:16:3 = 13.43, P <0.001), because the number of semi-erectoides segregates was significantly higher than expected. However, it appears that the deviation may have been due to chance since the segregation in the F3 was in accord with expectation (see later). One of the F2 plants from cross L197 x NGB5862 originally classified as semi-erectoides appeared different at the time of harvest and was re-classified as slender semi-erectoides (Fig. 1). It was longer in the early internodes than plants in the semi-erectoides class and had parthenocarpic pods of the inflated form seen in slender plants, rather than the flat parthenocarpic pods occasionally found in the ordinary lkb plants. In addition, the total height of the plant was greater than that of the semi-erectoides plants due to there being more nodes as a result of pod abortion. It was thought that this plant may be of the desired genotype (la cryS lkb) and this was confirmed by the F3 segregation (see later).
The F2 of cross L197 x NGB5865 consisted of 53 tall, 23 semi-erectoides, and 3 slender plants (Fig. 2). These numbers are in agreement with the expected 45 (La and/or Cry, Lka) : 16 (La and/or Cry, lka and la cryS lka) : 3 (la cryS Lka) ratio (c22 for 45:16:3 = 1.17, P>0.50). Unlike cross L197 x NGB5862, there were apparently no slender semi-erectoides (la cryS lka) segregates in the F2 generation, even though 80 seeds were planted.
Fig. 1. The stem length between nodes 1 and 5 plotted against the length between nodes 7 and 11 for each F2 segregate from cross L197 x NGB5862. The broken lines indicate the limits of the three classes - slender (la cryS Lkb), semi-erectoides (La and/or Cry, lkb) and tall (La and/or Cry, Lkb). The single plant lying to the right of the semi-erectoides group was shown in the F3 to be of the slender semi-erectoides phenotype (la cryS lkb).
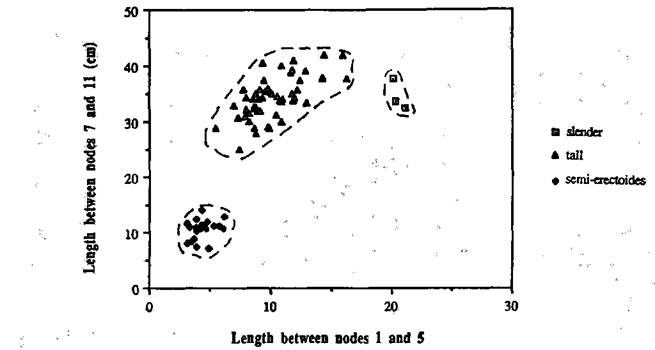
Fig. 2. The stem length between nodes 1 and 5 plotted against the length between nodes 7 and 11 for each F2 segregate from the cross L197 x NGB5865. The broken lines indicate the limits of the classes slender (la cryS Lka), tall (La and/or Cry, Lka) and semi-erectoides (La and/or Cry, lka).
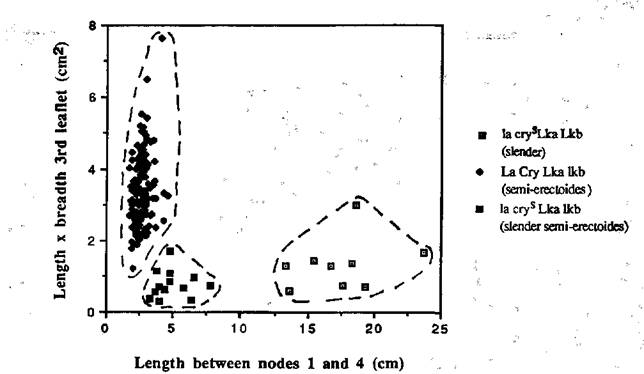
Fig. 3. The stem length between nodes 1 and 4 plotted against the length x breadth of the first leaflet at node 3 for each F3 segregate from selected slender and semi-erectoides F2 plants of cross L197 x NGB5862. The dotted lines denote the limits of the three classes - slender (la cryS Lkb), semi-erectoides (La and/or Cry, lkb) and slender semi-erectoides (la cryS lkb).
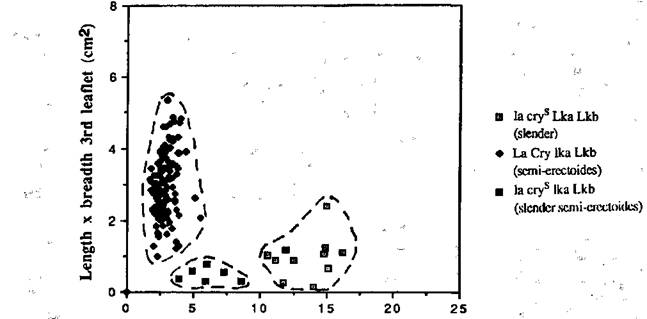
Fig. 4. The stem length between nodes 1 and 4 plotted against the length x breadth of the first leaflet at node 3 for each F3 segregate from selected slender and semi-erectoides F2 plants of cross L197 x NGB5865. The dotted lines denote the limits of the three classes -slender (la cryS Lka), semi-erectoides (La and/or Cry, lka) and slender semi-erectoides (la cryS lka).
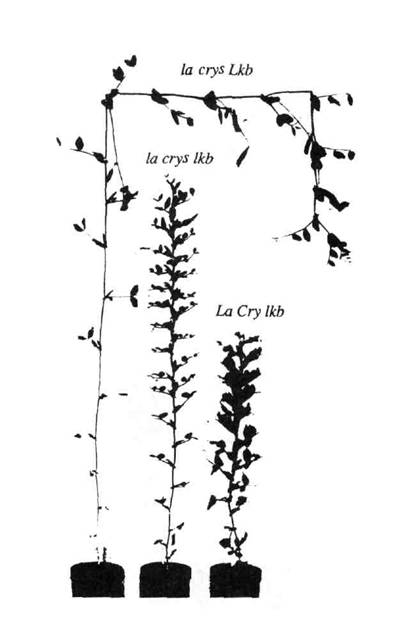
Fig. 5. The phenotype of a slender semi-erectoides plant (la cryS lkb, centre) with the parental lines from cross L197 (slender, la cryS Lkb, left) x NGB5862 (semi-erectoides, La Cry lkb, right).
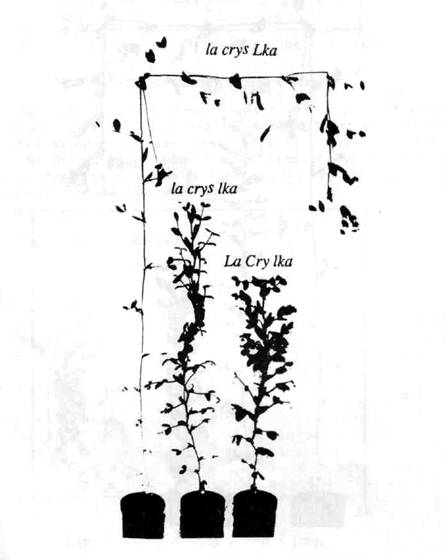
Fig. 6. The phenotype of a slender semi-erectoides plant (la cryS lka, centre) with the parental lines from cross L197 (slender, la cryS Lka, left) x NGB5865 (semi-erectoides, La Cry lka, right).
F3 progenies (normally 12 plants) were grown from 12 semi-erectoides and 3 slender plants in the F2 generation of cross L197 x NGB5862, and from 12 semi-erectoides and 2 slender plants in the F2 of cross L197 x NGB5865. In the F3 of both crosses, three distinct classes became apparent - slender (la cryS and Lka or Lkb), semi-erectoides (La and/or Cry and lka or lkb) and slender semi-erectoides (la cryS lka or lkb) (Figs 3 and 4). There were 5 slender semi-erectoides (la cryS lka) segregates in 3 progenies from semi-erectoides F2 plants and 2 in the 13 seeds obtained from slender F2 plants of cross L197 x NGB5865. The small numbers in the progenies from slender plants preclude the testing of statistical significance. However, the segregation in the progenies from semi-erectoides F2 plants is in accord with the expected 11:1 ratio (c21 = 3.48 with Yates' Correction, P > 0.05). In cross L197 x NGB5862 there were no slender semi-erectoides (la cryS lkb) segregates in the progenies of slender F2 plants, but 4 progenies from semi-erectoides plants segregated in the F3 generation and yielded 9 slender semi-erectoides(la cryS lkb) segregates. This is in accord with the expected 11:1 ratio for semi-erectoides (La and/or Cry, lkb) to slender semi-erectoides (la cryS lkb) types (c21 = 0.67).
The homozygous la cryS lka and la cryS lkb plants were phenotypically similar, and intermediate in height between their respective parental lines (Figs 5 and 6). They had longer internodes in the lower part of the plant than their semi-erectoides parent but the upper part of the plant was similar in length to this parent. Both also had the stem banding characteristics of the semi-erectoides parent (3). Both la cryS lka and la cryS lkb plants had a tendency to produce parthenocarpic pods similar to those produced by slender plants. The large number of parthenocarpic pods produced by the slender semi-erectoides plants meant that they had very low fertility. The leaves at the lower nodes of the slender semi-erectoides plants were similar to those of the slender parent, being small and narrow.
The similarity between la cryS lka and la cryS lkb plants raises the question of whether the phenotypes of NGB5862 (lkb) and NGB5865 (lka) have the same physiological and anatomical basis since the two mutants seem to interact in a similar way with the slender gene combination. However, anatomical studies suggest that this may not be the case (3). The phenotype of la cryS lka and la cryS lkb plants is effectively the superimposed effects of the slender (la cryS) combination and the semi-erectoides (lka or lkb) mutant. Consequently it is suggested that the intermediate stature of plants of genotypes la cryS lka and la cryS lkb reflects the fact that no direct relationships appear to exist between the slender and semi-erectoides genes in regard to the GA response process. There is some evidence that la_ and cryS are directly involved in the GA reception process (2) while it is possible that lka and lkb have no direct involvement in this process (3), possibly acting to increase the wall yield threshhold (1). One conclusion that can be drawn from these observations of the interaction of lka and lkb with the slender gene combination is that, unlike the GA-synthesis genes, le and na (2), both lka and lkb have the ability to reduce internode length on a slender background. It is hoped that the new genotypes, la cryS lka and la cryS lkb may provide valuable insights into the GA response process and the mechanisms underlying the various GA-sensitivity mutants in peas.
Behringer, F.J., D.J. Cosgrove, J.B. Reid, and P.J. Davies. 1990. Plant Physiol. 94:166-173.
Potts, W.C., J.B. Reid and I.C. Murfet. 1985. Physiol. Plant. 63:357-364.
Reid, J.B. and J.J. Ross. 1989. Physiol. Plant. 75:81-88.