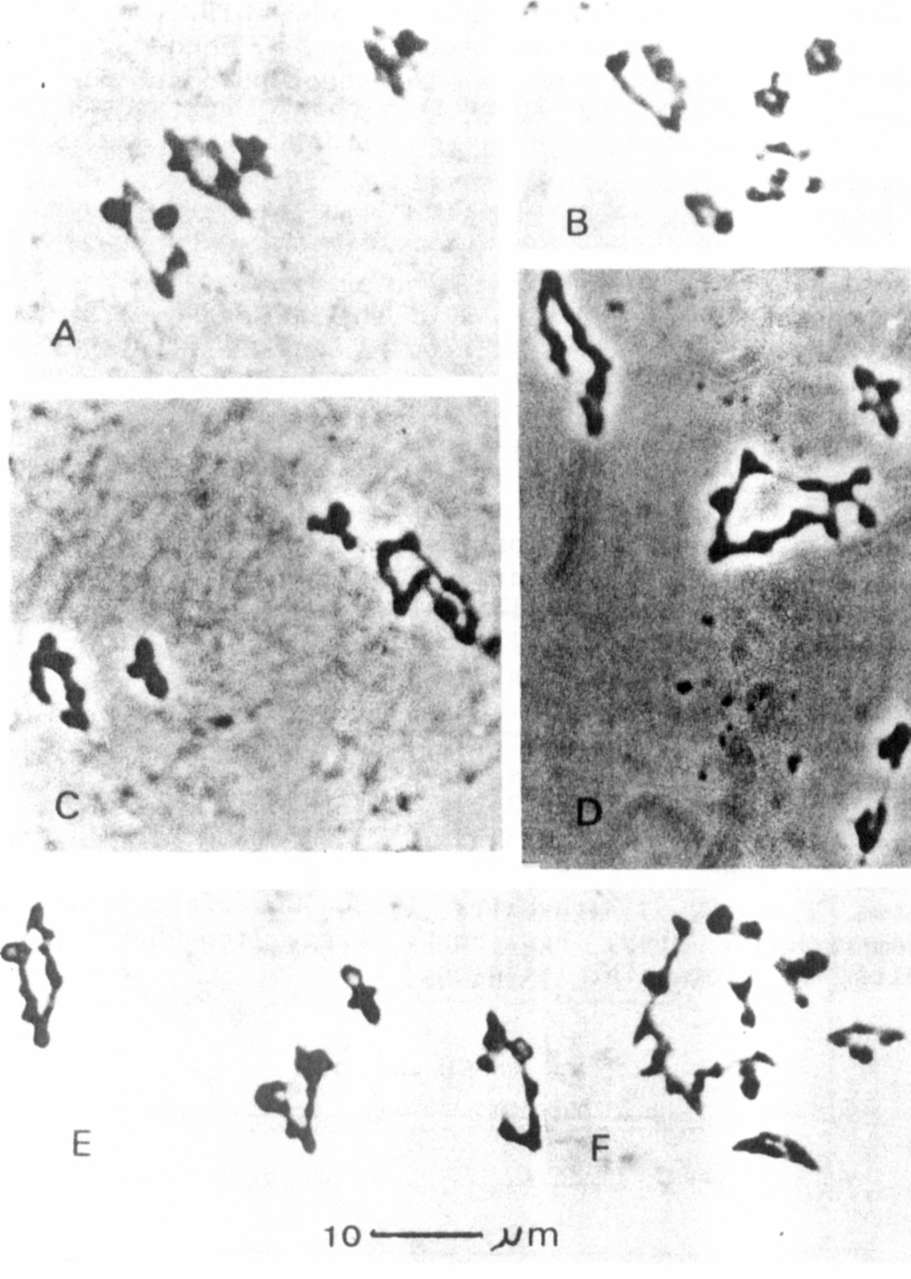
22
PNL Volume
19
1987
RESEARCH
REPORTS
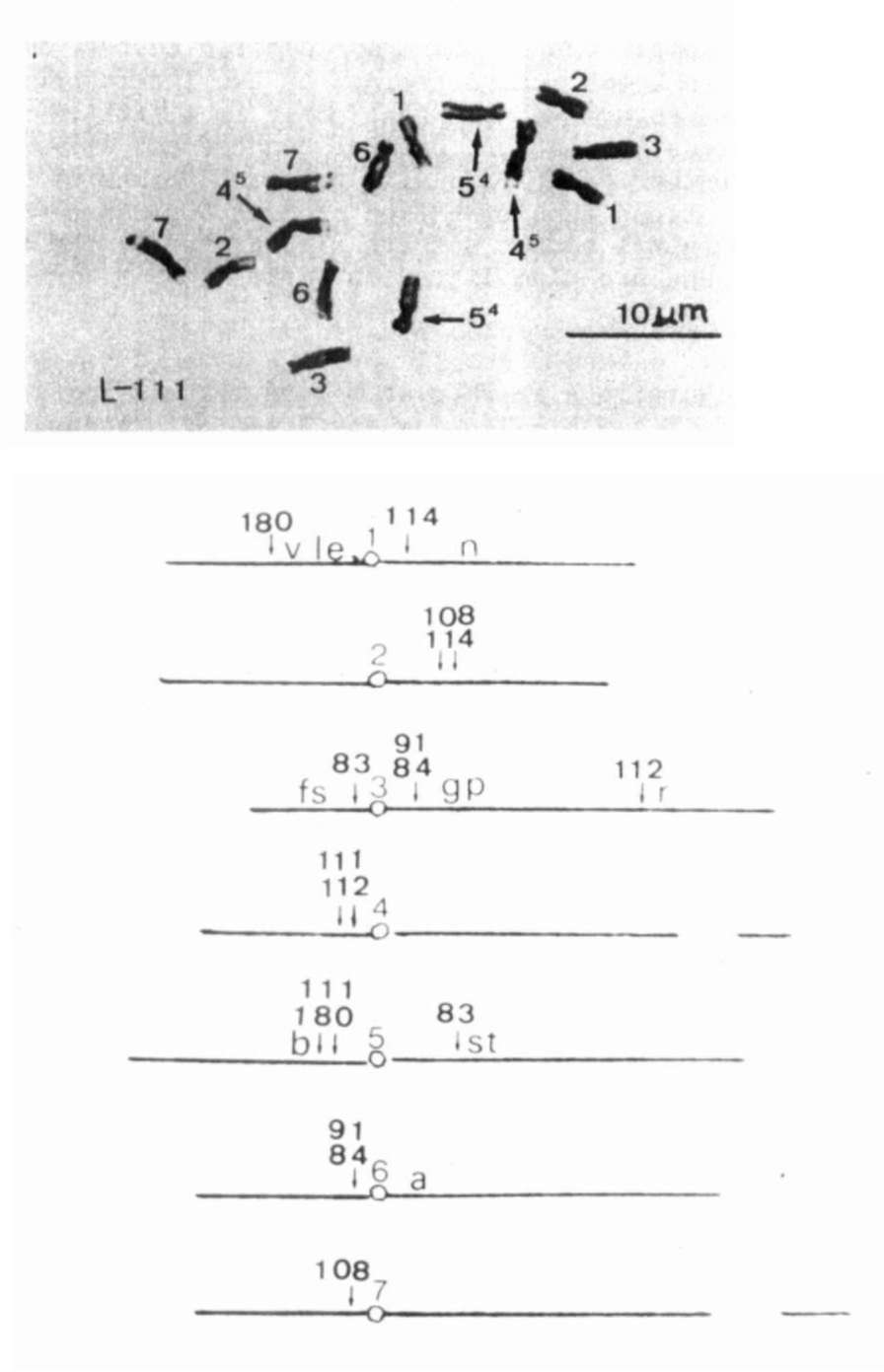
Fig. 1. At the top of this Fig.:
Metaphase oi mitosis in
a root tip of L-lll, Merton 1, T(4-5)a. Below:
Tentative chromosome maps with arrows indicating
the approximate positions of T-points in relation
to centromeres and neighboring marker genes.
a root tip of L-lll, Merton 1, T(4-5)a. Below:
Tentative chromosome maps with arrows indicating
the approximate positions of T-points in relation
to centromeres and neighboring marker genes.