PNL Volume 17 1985
RESEARCH REPORTS 87
Other investigators (1) confirmed these results and Przybylska ap-
pears to have detected a third allele at this locus in P. humile from
Jerusalem, described by her as a phenotype "c". A tight linkage between
the locus LAP-1 and the gene B on chromosome 3 was described for the
first time by Almgard and Ohland (1). Only two alleles at LAP-1 locus
have been detected in our investigations. We called them alleles A1 and
A2 (Fig. I) but they are not the subject of our interest in this paper.
The LAP-2 locus was mentioned by Scandalios and Espiritu (6) as
monomorphic. Polymorphism at this locus was described by Przybylska et
al. (3, 4) as well as by Gottschalk and Muller (2), who dealt with seed
protein analysis, and recently by Weeden and Marx (10).
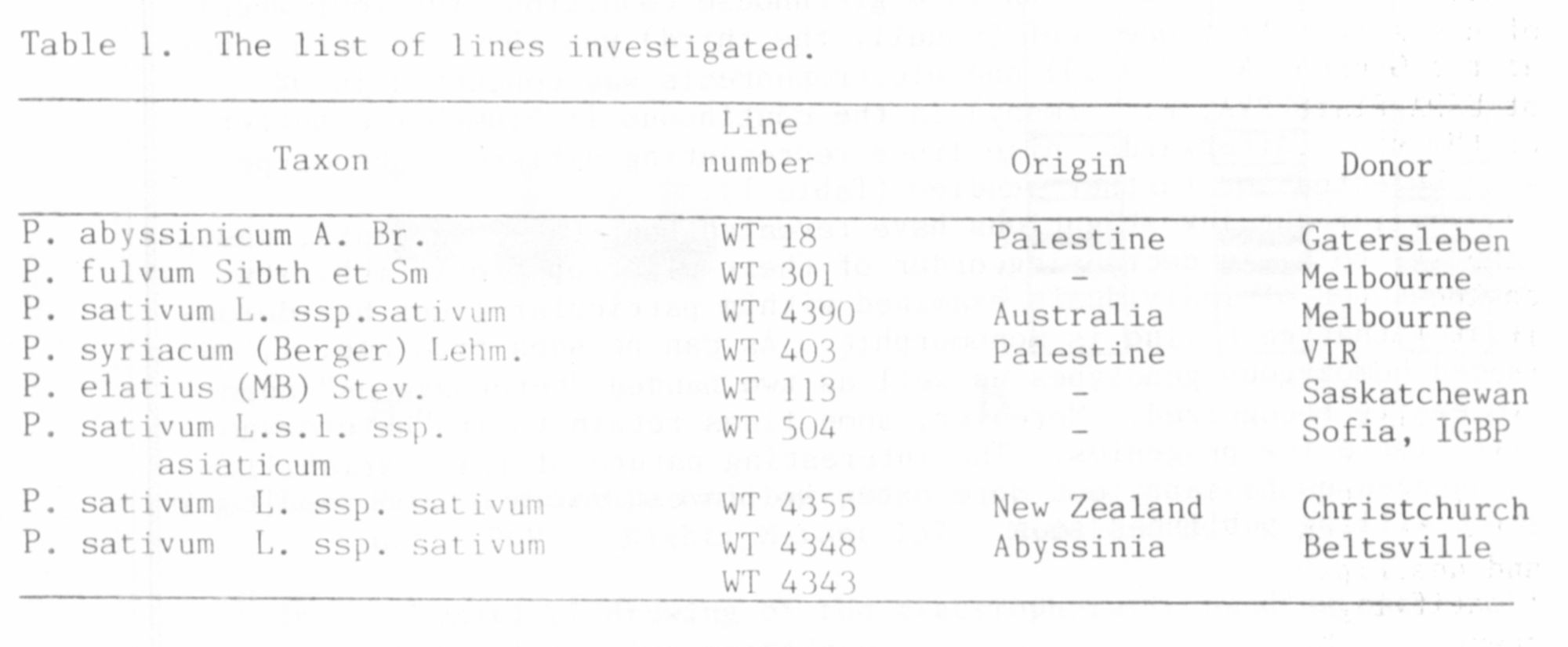
Ninety-five pea lines of the Wiatrowo Pea Gene Bank (8) belonging
to different Pisum taxa (P. sativum s.l. - 34 lines; P. abyssinicum - 8
lines; P. e1atius - 8 lines; P. fulvum - 3 lines; P. syriacum - 8 lines;
and 34 lines of the mutation group) were analyzed in this study. All
plants were cultivated in uniform greenhouse conditions for four weeks.
Crude extract from one leaf (usually the third) was absorbed onto paper
wicks (Beckman No. 319329) and electrophoresis was conducted in 9%
starch gel (SERVA, W. Germany) in the continuous lithium-boric buffer
system (5). Afterwards, nine lines representing different phenotypes
were selected for further studies (Table 1).
So tar our investigations have revealed four LAP-2 variants, desig-
nated a1 to a4 in decreasing order of their electrophoretic mobility. A
large number of individuals examined within particular lines has demon-
strated that each line is monomorphic. As can be seen from Fig. 1, one
banded homozygous genotypes as well as two banded "heterozygous" ones
are easily recognized. Moreover, some lines retain their "heterozygo-
sity" for a tew progenies. The interesting nature of this variability
encouraged us to carry out more extensive investigations, the results of
which will be published soon.

Fig. 1. Diagram of banding patterns in the Amp-1 and Amp-2 Pisum
forms: 1-Wt 18, 1-Wt301, 3-Wt 4390, 4-Wt 403, 5-Wt 113,
6-Wt 504, 7-Wt 4355, 8-Wt 4348, 9-Wt 4343.