MORPHOLOGICAL VARIATION IN PLANTS REGENERATED FROM LONG-TERM CALLUS
CULTURE OF PEA
Ezhova, T. A., A. M. Bagrova, and S. A. Gostimski
Moscow State University, USSR
Pea callus tissue was derived from apices of var. 'Ranny zeleny-33'
using the method of Gamborg, et al. (1). In these conditions a friable,
light-green, rapidly growing morphogenetic callus was obtained. The
callus was transferred to fresh medium every 1-1/2 to 2 months over a
two-year period. To secure a large number of shoots, two-year-old
callus was placed into 250 ml Erlenmeyer flasks either on the same
medium or on a medium with added benzylaminopurine content (2 mg/1). To
avoid getting adventitious shoots from the same meristematic zones, 3-5
cm long shoots were taken from flasks along with the pieces of callus
from which they had been derived.
Shoot tips were grafted in the greenhouse. From about 200 shoots
grafted, seeds were obtained from 70 Regenerants (Ro).
Some of the grafted regenerants had shortened internodes and
altered leaf and flower morphology.
Seeds from the grafted regenerants (from 1 to 15 seeds from each
regenerant) were planted in the field. Segregation of a chlorotica
mutant was observed in one of 65 lines (4 green : 1 chi plants).
Another line segrated for a waxless mutant (2 normal : 1 waxy plants).
It is likely that other mutations had occurred but , because the popula-
tion sizes were small, they remained undetected. Therefore analysis of
the R2 generation is required.
The most interesting results were obtained while studying some
physiological and quantitative characters. In 70-80% of the R1 Lines
the following changes were found: more robust habit as compared with the
initial variety (IL); dark green leaves unlike the light green leaves of
the IL; oblong leaflets as compared with oval; earlier or later flo-
wering time.
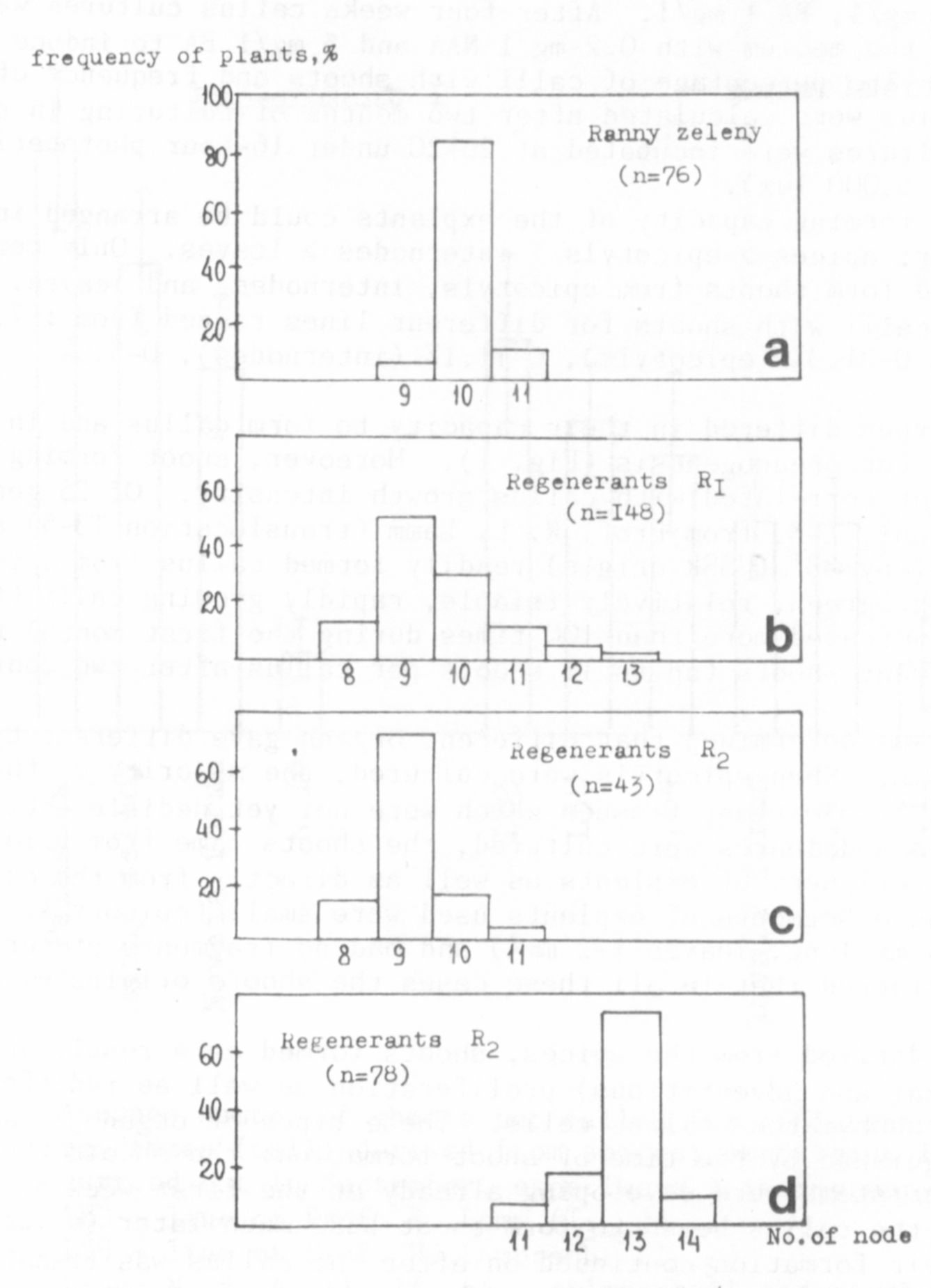
For example, 85.5% of the plants of var. Ranny zeleny formed theit
first flower on node 10 (14.5% on nodes 9 and 11). Among the R1 regene-
rants only 27% of the plants flowered at node 10, while the rest (73%)
flowered at nodes 8, 9, 11, 12 and 13 (Fig. la, b).
We have not seen direct correlations among these changed charac
ters. For instance, some plants with more vigorous habit were late flo-
wering lines and some were early.
The same changes that occurred in R1 also were present in R2 and
there was no segregation of the changed characters among the plants and
the families belonging to the same line. For example, all the plant
among three families of line No. 1 had statistically significant differ-
ences (P > 0.999) in the length/width ratio of the leaflets. Thus the
change in leaflet shape (from oval in the IL to oblong) was an inherited
difference (Table 1).
The study of flowering time of nine lines of the R2, general ion has
shown that five lines flowered more frequently on node 9 (from 8 to 10,
very rarely on node 11), and four lines flowered at node l3 (from 11 to
14) (Fig. 1 c, d). These data indicate that variability of flowering
time among R1 lines did not result from physiological and epigenetical
events, but had genetic basis.
Studies of karyotypes in callus tissues, pollen fertility, and
meiosis in regenerants, which is to be reported, demonstrated that the
variability of regenerants was not caused by changes in ploidy or by
large chromosomal aberrations.