40
PNL Volume 17
1985
RESEARCH REPORTS
DETAILED DOCUMENTATION OF THE OBSCURATUM PHENOMENON IN PISUM
Loennig, W.-E.
Institute of Genetics, University of Bonn
Federal Republic of Germany
In a line of Pisum sativum (arvense) there appeared the phenomenon
long known as obscuratum (1, 3-7, 9), showing a certain percentage of
self-colored, violet-black seeds among a large majority of violet
spotted ones (F, Fs, or F Fs). The character requires the presence of
anthocyanin (A) for expression but it is not heritable. Although the
phenomenon has often been observed and described, few data are available
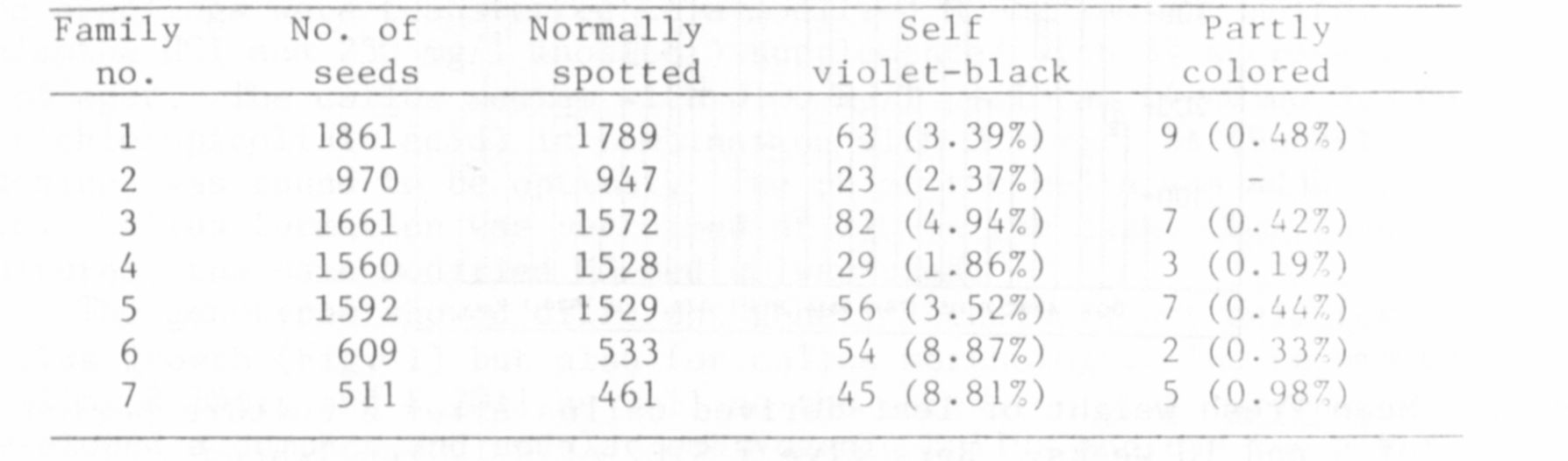
providing actual counts and percentages. Tables 1 and 2 give such data
gathered in the experimental fields of Bonn in 1983 and 1984. Table 1
shows the numbers of seeds and the percentages of violet-black seeds in
seven families derived from individual plants which themselves bore only
spotted seeds.
Table 1. Percentages of violet-black and partly colored seeds of
seven families of a line of Pisum arvense (1983).
The violet-black seeds from these populations were selected and the 184
plants grown from them were investigated in the field in 1984 (Table 2).
Table 2. Percentages of violet-black and partly colored seeds. All
the plants producing these seeds were grown from violet-black
seeds.

The 122 self-colored or partly colored seeds were distributed on 24
(13%) of the 184 plants; all 24 plants also bore spotted seeds. Three
categories could be distinguished for the colored seeds: five plants
bore only violet-black seeds, eight plants bore violet-black and partly
colored seeds, and eleven plants had only partly colored seeds, so that
the ratio of the first two categories to the last was nearly 1:1 (in
four groups of plants we found the following ratios: 6:6 in 71 plants;
2:2 in 48 plants; 3:2 in 44 plants; and 2:1 in 21 plants).

